银屑病发病机制的研究进展
时间:2023-08-06 11:31:03 热度:37.1℃ 作者:网络
银屑病是一种由免疫介导的多基因调控的慢性炎症性疾病,发病率为2%~3%。银屑病临床治疗难度较大,且容易复发,严重危害患者身心健康,影响患者生活质量,对于其发病机制的研究,一直以来均是皮肤科研究的重点和难点。
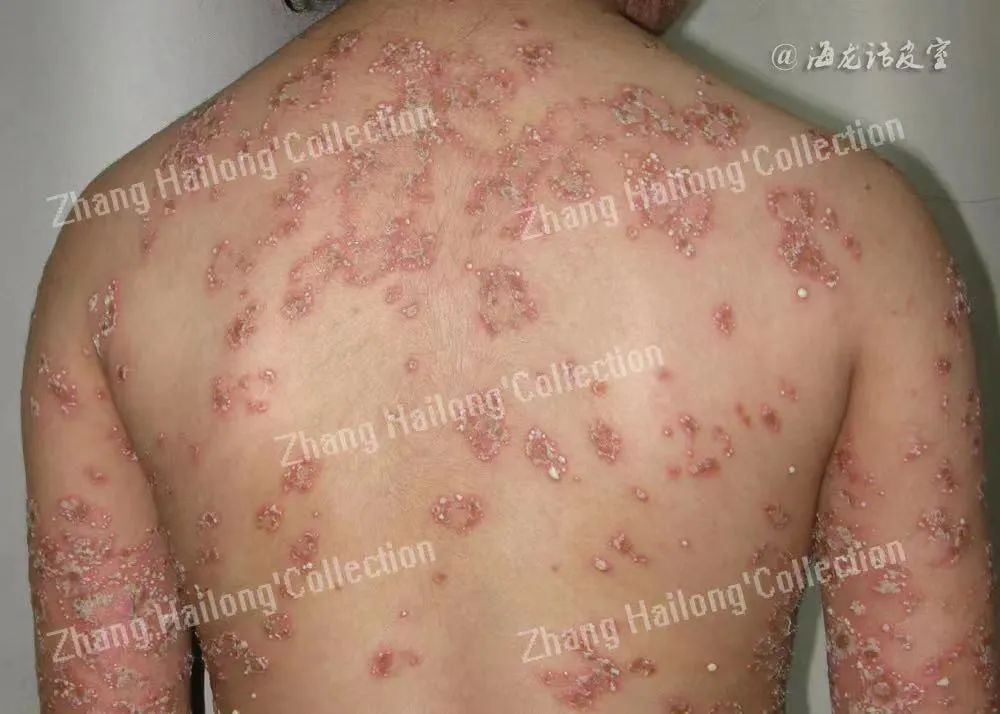
银屑病的病因复杂,其发生发展可能涉及T细胞分化、炎症细胞浸润、角质形成细胞增殖等。此外,遗传、感染、代谢障碍、内分泌异常及其他相关因素(吸烟、饮酒、外伤、劳累、精神因素等)也可诱发甚至加重银屑病。 目前已在遗传、免疫等领域对银屑病的发病机制进行了大量深入的研究,但其发病机制尚未完全阐明。
近期关于银屑病发病机制的研究进展提高了人们对该疾病机制的认识,并推动了免疫生物学和小分子学科的发展,有助于控制其相关慢性炎症。
角质形成细胞的生物学异常
银屑病是一种由免疫机制介导、多基因遗传调控的炎症病变,参与其皮损部位免疫反应的细胞主要包括角质形成细胞、朗格汉斯细胞、淋巴细胞、抗原呈递细胞等,其组织病理学表现为角质形成细胞角化过度伴角化不全,炎症细胞浸润及毛细血管增生、扩张。异常角质形成细胞作为银屑病致病细胞的概念已得到人们的广泛关注。JunB/c-Jun异常表达或角质形成细胞中信号转导及转录激活因子(signal transduction and activator of transcription,STAT)3磷酸化小鼠发生的皮肤炎症反应具有银屑病的组织学和分子学特征。注射辅助性T细胞(helper T cell,Th细胞)17相关细胞因子(如IL-23或IL-21)小鼠的皮肤可发生具有银屑病形态学特征的表皮增生,并伴有炎性T细胞浸润。同时,角质形成细胞本身也可生成细胞因子,其 中主要成员为IL-8一种最初在银屑病鳞屑中发现的细胞因子。细胞因子IL-15在银屑病表皮中表达,保护角质形成细胞不凋亡。此外,可溶性IL-15受体α也可通过抑制角质形成细胞分泌细胞因子并抑制生成IL-17的T细胞的扩增,从而有效控制银屑病的发生。角质形成细胞是IL-1的主要生成来源。一些细胞因子可触发炎症小体活化和角质形成细胞分泌IL-1,而导致银屑病炎症反应的发生。其他与银屑病发病机制相关的由角质形成细胞产生的介质包括抗菌肽、S100A8/9、β-defensins及抗菌肽(LL-37)。综上所述,影响角质形成细胞的转录因子可促使银屑病的发生。
免疫细胞的重要作用
目前,银屑病的免疫学发病机制是银屑病研究的热点,银屑病的发生和发展与免疫细胞密切相关。最初用于预防器官移植排斥反应的免疫抑制剂在银屑病的临床治疗中发挥了极其重要的作用。随后,免疫抑制剂(如氨甲蝶呤或环孢霉素)开始广泛应用于各种类型银屑病的治疗。人白细胞抗原的相关性遗传数据以及破损皮肤存在的寡克隆T细胞及其对皮肤抗原的反应性数据,进一步强调了免疫细胞在银屑病发病中的重要性。组织中存在着重要的记忆性T细胞群,其参与了局部免疫应答,这些特定的记忆T细胞被命名为组织常驻记忆T细胞(tissue resident memory T cell,TRM)。TRM主要存在于呼吸道、生殖道和皮肤等上皮屏障组织中。当体内发生感染时,TRM可迅速做出反应,控制住病原体的扩散,因此TRM对保护宿主免受有害微生物侵袭起至关重要的作用。研究显示,银屑病患者局部皮肤上皮组织中的TRM增加。
Th17细胞免疫应答与基因多态性
与银屑病相关的基因多态性包括基因编码的IL-23A、IL-23 受体、STAT3、Runt相关转录因子3和酪氨酸激酶(tyrosine kinase,TYK)2,上述基因均与Th17细胞免疫应答相关。Th17细胞的分化及活化取决于细胞因子IL-6、IL-21、IL-1、转化生长因子-β及IL-23。而IL-23及其受体和下游信号分子STAT3均与银屑病的遗传易感性有关。此外,STAT3可被IL-6和IL-21激活, 并与其他Th17细胞共同激活转录因子维甲酸相关孤儿受体γt,其中STAT3主要表达IL-17A和IL-17F。Th17细胞可能是银屑病发病机制的核心调控因子,其可与角质形成细胞、内皮细胞以及多种免疫细胞相互作用。记忆Th17细胞的重新激活可能是导致银屑病慢性病程的主要原因。
相关细胞因子水平的异常升高
如何针对患者具体的病情特点,更好地配合运用中医和西医,制定个体化治疗方案,使其达到最优效果,是现阶段我们仍需努力探索的。IL-17A及IL-22由固有淋巴细胞(innate lymphoid cells,ILCs)和γδT细胞的免疫细胞生成。ILCs作为一种独特的先天免疫细胞群,缺乏抗原特异性受体,可被细胞因子刺激产生一系列效应细胞因子。目前,ILCs根据产生细胞因子和特定转录因子的特性分为ILC1、ILC2、ILC3。在 ILCs群体中,包括淋巴组织诱导细胞在内的ILC3的特点是伴随维甲酸相关孤儿受体 γt 的高表达,生成IL-17A和(或)IL-22。以人群为例,ILC3可根据自然杀伤细胞(natural killer cell, NK细胞)p46的表达模式分为NKp46+及NKp46-两个亚群。由于银屑病患者外周血和皮肤中IL-17A和IL-22产生的NKp46+ILC3水平均升高,故推测NKp46+ILC3参与了银屑病的发病。维甲酸相关孤儿受体γt+CD56+ILC3的发现证实,ILC3亚群在银屑病发病中起关键作用,能够使银屑病患者的皮肤内高度蓄积 IL-22。皮肤内IL-17A的细胞来源是γδT细胞群体。大多数的真皮γδT细胞表达的T细胞受体包含Vγ4及CC趋化因子受体6。使用Toll样受体7激动剂咪喹莫特的银屑病炎症小鼠模型皮肤的真皮Vγ4+Vδ4+T细胞生成IL-17A和IL-17F,导致皮肤炎症。有报道证实,银屑病患者体内的肥大细胞可以产生IL-17A和IL-22。值得注意的是,现已发现的与银屑病发病及治疗相关的免疫细胞均可产生TNF。
综上所述,银屑病的发病机制复杂,涉及免疫应答、炎症介质、细胞因子、基因遗传等因素。同时,与环境因素息息相关,如吸烟、饮酒及精神压力 等均可诱发或加重银屑病。
参考文献:
[1]Korman NJ.Management of psoriasis as a systemic disease: What is the evidence?[J].Br J Dermatol,2020,182(4) : 840-848.
[2]Chiricozzi A,Romanelli P,Volpe E,et al.Scanning the Immuno-pathogenesis of Psoriasis[J].Int J Mol Sci,2018,19(1) : 179.
[3]Boehncke WH,Brembilla NC. Unmet Needs in the Field of Pso-riasis: Pathogenesis and Treatment[J].Clin Rev Allergy Immu-nol,2018,55(3) : 295-311.
[4]Bouchaud G,Gehrke S,Krieg C,et al. Epidermal IL-15Rα acts as an endogenous antagonist of psoriasiform inflammation in mouse and man[J].J Exp Med,2013,210( 10) : 2105-2117.
[5]Gross O,Yazdi AS,Thomas CJ,et al. Inflammasome activators induce interleukin-1α secretion via distinct pathways with differen-tial requirement for the protease function of caspase-1[J].Immu-nity,2012,36(3) : 388-400.
更多参考文献略。。。

