癌症生物学系列:癌症中的生长因子与受体
时间:2024-06-04 21:01:05 热度:37.1℃ 作者:网络
1 前言
癌基因及其前体——原癌基因的发现促使人们提出了一系列问题,其中居于核心地位的问题是,癌基因到底如何通过它们所编码的蛋白质成功地扰乱了细胞的行为?
对这个问题的理解,首先要了解正常细胞的生命活动。细胞接收并处理生化信号从而调节细胞增殖,正常的癌基因编码蛋白质常常是这一机制的组成部分。正常组织形态维持的实现皆有赖于细胞间的协同合作,而大部分对话是由生长因子负责传达的。这些相对分子量较小的蛋白经细胞分泌,于细胞间隙运行,最终作用于其他细胞的受体,传递它们独特的生物学信息。
在癌症中,使细胞感知周围环境中生长因子的存在、将信号传入细胞内部,并进一步处理这些信号的信号分子被癌基因编码蛋白质篡权。通过取代正常细胞的生长刺激机制,癌蛋白可诱使细胞相信周围环境中存在的正是生长所需的生长因子。一旦诱导成功,细胞便会开始增殖,其反应与大量生长因子存在于其周围介质所介导的反应一样。
2 癌蛋白和生长因子受体的酪氨酸激酶功能
细胞间信号如何通过生长因子传递的第一条线索来源于对癌基因v-src及其特定蛋白质产物的生化分析。1978年,生化学家发现了Src发挥作用的机制。他们研制出能与Src特异识别并结合的抗体,当其在同时含有Src及腺苷三磷酸的溶液中孵育时,该抗体被磷酸化。由此可知,Src是作为蛋白激酶发挥作用的,即将ATP的一个高能磷酸键转移至一个适宜的蛋白质底物。
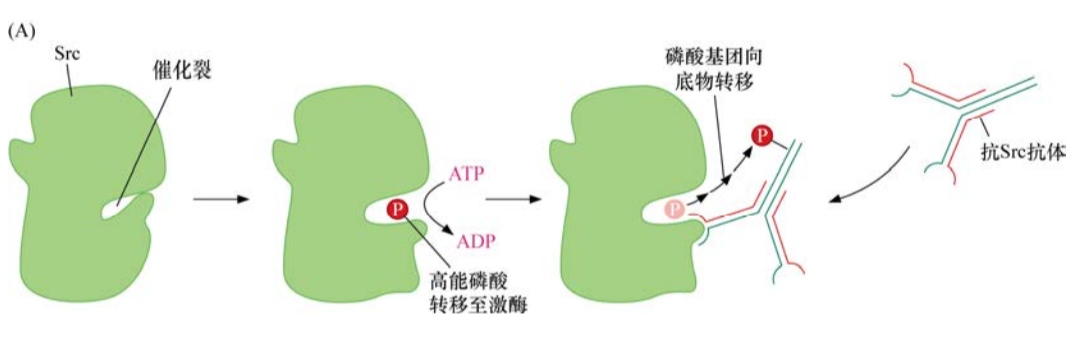
Src激酶功能的揭示意义深远。原则上,一个蛋白激酶可以磷酸化细胞内多种不同的蛋白质底物。一旦被磷酸化,这些底物蛋白质的功能可能会发生改变,进而改变其下游靶点的功能。这种作用机制似乎可以解释像Src这样的蛋白为何能如此多效地干扰多种细胞的表型。
不久之后,人们又发现Src与以前发现的其他蛋白激酶显著不同。已知其他的蛋白激酶是将磷酸基团连接到丝氨酸和苏氨酸残基的侧链上。而然Src则磷酸化蛋白质底物的酪氨酸残基。在正常细胞内99%以上的磷酸化氨基酸为磷酸化丝氨酸和磷酸化苏氨酸,磷酸化酪氨酸仅占这些细胞全部磷酸化氨基酸的0.05%-0.1%。
在随后的研究中人们得出了另一条与之相关的结论:在哺乳动物细胞中,通过酪氨酸磷酸化传递信号的这种机制主要出现在丝裂原信号转导通路中,而其他信号转导途径中所涉及的成千上万的激酶,则基本上全部依赖于磷酸化丝氨酸和磷酸化苏氨酸来传递信息。那么酪氨酸残基的磷酸化是如何导致细胞转化的呢?
对这一问题的回答要提到上皮生长因子——EGF。人们发现,EGF对多种上皮细胞具有促有丝分裂作用。EGF通过作用于一种细胞表明蛋白——EGF受体(EGFR),并与之结合,将信息向胞内传递。对EGFR的整体结构特征分析,发现EGFR分为3个主要区域,较大的N端胞外结构域,主要负责识别并结合EGF;一段23个氨基酸的跨膜区,这段疏水氨基酸构成区域可以在细胞膜的脂质双层中稳定存在;最后,C端由542个氨基酸构成的第三个结构域,伸展进入细胞质。对胞内结构域的检测显示,它与已知的Src蛋白质序列有明确的序列相似性。
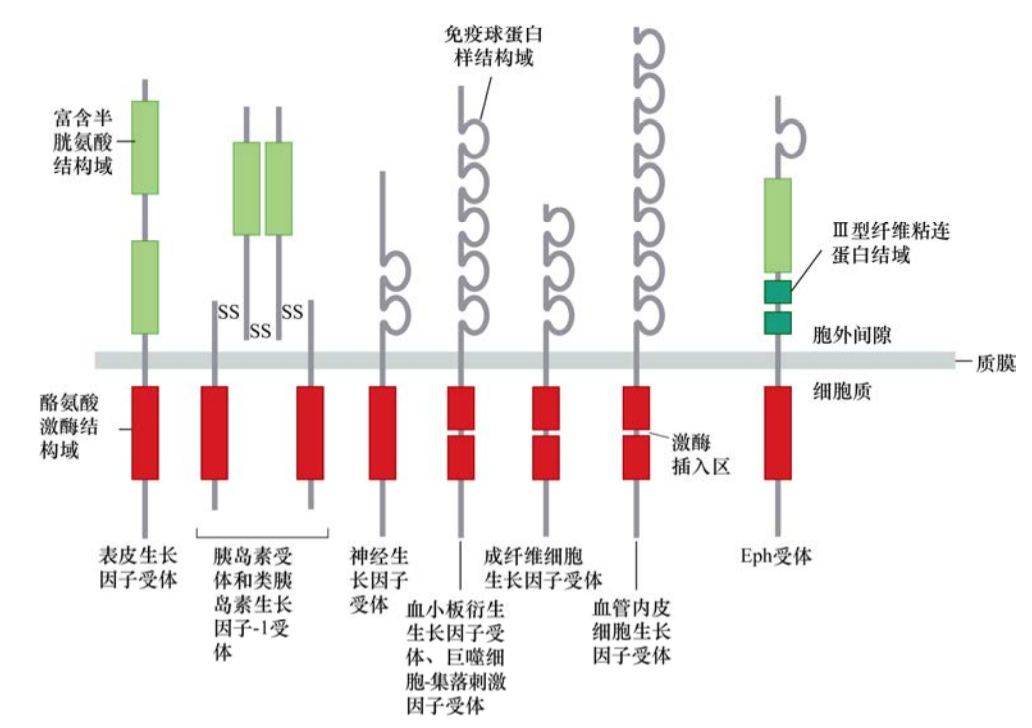
这样,答案瞬间变得清晰起来:一旦EGFR结合EGF,其胞内结构域的Src样激酶将以某种方式被激活,之后磷酸化某些胞内蛋白质的酪氨酸残基,从而使细胞增殖。随后的测序试验结果显示,许多酪氨酸激酶都有整体的结构相似性,其中许多可产生癌蛋白样作用。
3 被改变的生长因子受体可发挥癌蛋白样作用
1984年,人们证实EGF受体序列与已知的由erbB癌基因编码的癌蛋白序列高度相关,这一发现极具震撼力。由此,细胞生物学的两个不同领域被联系起来,一个被细胞用于感知周围环境中生长因子的蛋白质被挪用为一种由逆转录病毒基因编码的强力癌蛋白。
一次具体的研究中,人们发现erbB癌基因编码的癌蛋白缺少EGF受体N端胞外结构域的序列。在没有N端序列的情况下,ErbB癌蛋白无法辨认及结合EGF,然而,它发挥了细胞增殖刺激因子的功效。这一发现引发了一个有趣的推论并随后被证实:胞外结构域的缺失可通过某种方式使被截短的EGF受体蛋白向细胞内源源不断地发送生长刺激信号,而完全不依赖于EGF。多年后,人们在1/3的人成胶质细胞瘤中发现了这些被截短的EGF受体。而后,在乳腺癌中发现了ErbB/EGF-R的同源分子,如ErbB2、HER2及Neu,它们与不良预后密切相关。
ErbB-EGFR之间的关系为癌细胞的特性提供了一个简单明了的解释:ErbB癌蛋白所释放的信号与那些经配体激活的EGF受体所释放的信号非常相似。然而,不同于EGFR的是,ErbB癌蛋白可以持续不断地向细胞内发放生长刺激信号,从而使细胞认为周围存在着大量的EGF,而其实那里可能什么都没有。
4 生长因子基因可转变为癌基因
1983年,人们发现PDGF的B链序列与由猴肉瘤病毒v-sis癌基因编码的癌蛋白高度相关。PDGF与sis-编码癌蛋白间的联系向我们提供了另一个癌蛋白转化细胞的重要机制:猴肉瘤病毒感染细胞后,它的sis癌基因促使被感染细胞释放大量PDGF样sis蛋白到其周围细胞外空间中。在那里,PDGF样分子与释放它们的自身细胞上的PDGFR结合。其结果是造成这些细胞上PDGF受体强烈活化,在大量细胞中该配体激活PDGFR产生持续的生长刺激信号。
有丝分裂信号体系中的蛋白质又一次与病毒癌蛋白紧密地联系在了一起。病毒感染细胞合成并释放刺激自身产生应答的生长因子,而并非由一种类型的细胞向其附近的另一种类型的细胞发送生长因子信号(旁分泌),或由机体中的一种组织经由循环系统向另一远隔组织发放信号(内分泌)。它代表的是一种自我刺激,或自分泌信号系统,该系统中的细胞可生产作用于自身的有丝分裂原。
事实上,我们已知许多肿瘤细胞可以产生并释放大量它们自身亦可应答的生长因子。
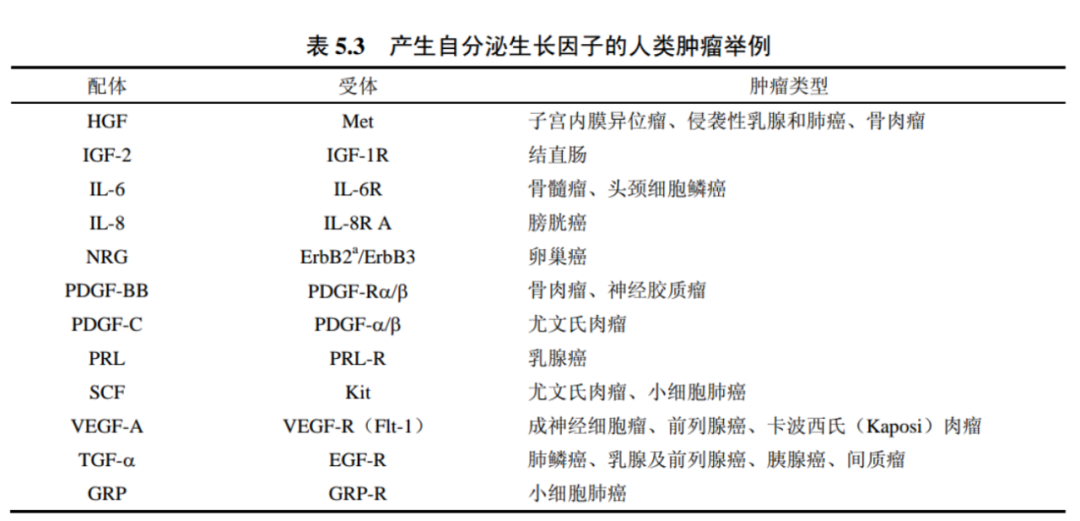
5 受体酪氨酸激酶依赖磷酸化发挥作用
通过为细胞提供持续生长信号刺激,癌蛋白能够驱动周而复始的细胞生长、分裂,那么具有酪氨酸激酶活性的生长因子受体是如何成功地将信号从胞外传递到胞内的呢?
为了解决这个问题,研究者运用EGF生长因子处理能够表达其同源受体EGFR的细胞,几秒钟后检测蛋白的磷酸化。结果显示,伴随配体与生长因子受体的结合,多种胞质蛋白的酪氨酸残基发生了磷酸化。这些蛋白中发生酪氨酸磷酸化最显著的是受体分子本身,因此,这些受体看起来是能够自磷酸化的。
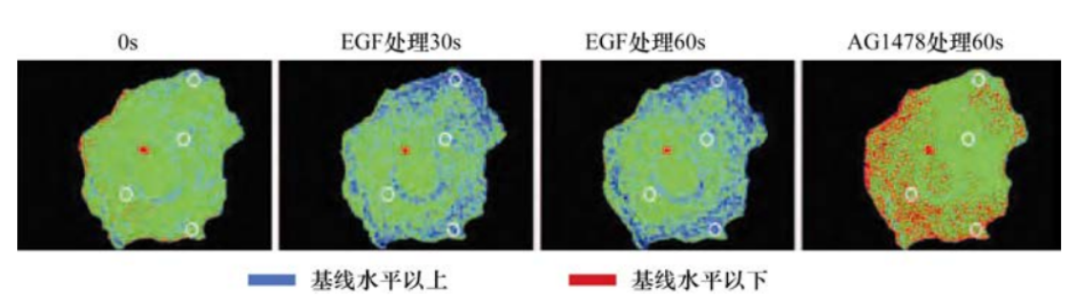
另一个线索来源于很多生长因子配体的结构:它们通常是二聚体,形成同源二聚体或异源二聚体。第三条线索来源于很多跨膜蛋白有类似EGF和PDGF受体的结构,在质膜上具有横向流动性。这些线索提供了一个简单的模型,当没有配体的情况下,一个生长因子受体通常以单体的形式嵌入质膜。当其生长因子配体存在时,受体分子就会结合在配体两个亚基的一个上。其后,配体-受体复合物将在质膜上徘徊直到有第二个机会遇到另一个受体分子,配体中没有结合受体的那个亚基将和第二个受体分子结合。这样,通过二聚体形式的配体为桥梁,把两个受体分子成功地交联在一起。
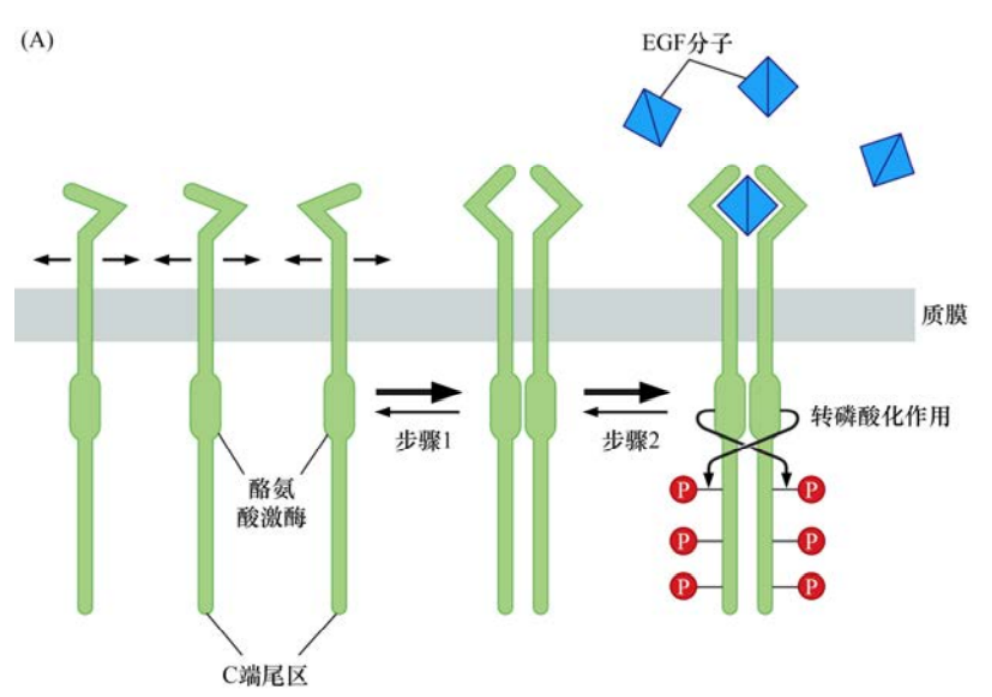
一旦两个受体分子的胞外结构域通过与配体结合而发生二聚化,胞质部分通常也会被拉在一起。每个激酶结构域磷酸化另一个受体胞质结构域里的酪氨酸残基。这种双向的相互磷酸化作用被称为转磷酸作用。
这些酪氨酸残基的磷酸化至少导致两种后果。激酶催化裂隙(即蛋白酶发挥功能的区域)通常受蛋白质的环结构阻隔,从而阻止激酶与其底物有效结合。阻隔“活化环”的关键酪氨酸的转磷酸作用可导致该环结构移开,从而可使底物分子直接进入催化裂隙。另外,转磷酸作用还可导致生长因子受体胞质部分激酶区域以外的一系列酪氨酸残基的磷酸化。这些磷酸化事件可使受体分子激活多种下游信号通路。
参考文献:
1. 《The biology of CANCER》second edition. Robert.A Weinberg

