【论著】| SIRT3通过去乙酰化YME1L1诱导乳腺癌内分泌治疗耐药的作用机制研究
时间:2024-07-21 06:00:34 热度:37.1℃ 作者:网络
[摘要] 目的:沉默调节蛋白家族(sirtuins,SIRT)是一类以烟酰胺腺嘌呤二核苷酸(NAD+)为辅酶的第Ⅲ类组蛋白去乙酰化酶。YME1样三磷酸腺苷酶(YME1 like 1 ATPase,YME1L1)对于维持线粒体形态、功能和可塑性至关重要。视神经萎缩相关蛋白 1(optic atrophy 1,OPA1)主要介导线粒体融合。本研究拟探索乳腺癌内分泌治疗耐药中SIRT3的表达变化, SIRT3与YME1L1、OPA1之间的关系及在乳腺癌内分泌治疗耐药中的作用机制。方法:使用4-羟基他莫昔芬(4-hydroxytamoxifen)诱导耐他莫昔芬(tamoxifen,TAM)的MCF-7/TAM。采用细胞计数试剂盒(cell counting kit-8,CCK-8)检测细胞增殖能力,验证耐药性。采用透射电镜和免疫荧光染色(immunofluorescence staining,IF)实验观察线粒体形态。通过实时荧光定量聚合酶链反应(real-time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)、蛋白质印迹法(Western blot)检测SIRT3、OPA1的基因表达和蛋白水平。采用JC-1染色检测线粒体膜电位,采用二氢乙啶(dihydroethidium,DHE)染色检测活性氧,验证线粒体功能。采用RNA干扰技术在耐药细胞中敲低SIRT3,采用过表达质粒在亲本细胞中过表达SIRT3及YME1L1基因野生型(wild type,WT)、模拟乙酰化状态突变型(mutant,MUT K237Q)、模拟去乙酰化状态突变型(MUT K237R)。采用免疫沉淀技术(immunoprecipitation assay,IP)及IF分析SIRT3与YME1L1之间的相互作用。结果:RTFQ-PCR及Western blot检测结果显示,SIRT3在耐药细胞中的表达显著高于亲本细胞。在亲本细胞中过表达SIRT3,乳腺癌细胞对TAM的敏感性出现下降。在耐药细胞中敲低SIRT3,耐药细胞对TAM的敏感性增强。DHE染色结果显示,在相同浓度的TAM处理下,耐药细胞中ROS水平低于亲本细胞;透射电镜及IF结果显示,相较于亲本细胞,耐药细胞的线粒体伸长;Western blot检测结果显示,耐药细胞L-OPA1蛋白表达水平高于亲本细胞。在亲本细胞中过表达SIRT3,线粒体功能增强,其形态较对照组更长;此外,L-OPA1表达上调;而在耐药细胞中敲低SIRT3后,得到相反结果。为了进一步验证SIRT3如何调控OPA1蛋白,影响线粒体的形态及功能,促进乳腺癌耐药,我们在亲本细胞中过表达YME1L1(野生型及突变型质粒),结果显示,过表达模拟去乙酰化状态的YME1L1与过表达SIRT3结果相似,过表达乙酰化状态的YME1L1得到与敲低SIRT3相似的结果。IP实验证实,SIRT3与YME1L1在乳腺癌细胞中存在相互作用;在SIRT3不同表达水平下,YME1L1乙酰化水平不同。IF实验显示,在MCF-7细胞中YME1L1与SIRT3存在共定位。结论:SIRT3在耐TAM的乳腺癌细胞中高表达。SIRT3通过去乙酰化YME1L1上调L-OPA1表达,进而促使线粒体融合并增强线粒体功能,促进乳腺癌对TAM耐药。
[关键词] 乳腺癌;内分泌耐药;SIRT3;YME1L1;OPA1
[Abstract] Background and purpose: Silent information regulator proteins (sirtuins, SIRT) are a class Ⅲ histone deacetylases with nicotinamide adenine dinucleotide (NAD+) as coenzyme. YME1 like 1 ATPase (YME1L1) is essential for the maintenance of mitochondrial morphology, function and plasticity. Optic atrophy 1 (OPA1) mainly mediates mitochondrial fusion. The aim of this study was to explore the expression of SIRT3 in the endocrine resistance of breast cancer, the relationship between SIRT3 and YME1L1 and OPA1, and the mechanism of SIRT3 in the endocrine resistance of breast cancer. Methods: 4-hydroxytamoxifen was used to induce tamoxifen-resistant MCF-7/TAM cells. cell counting kit-8 (CCK-8) was used to detect cell proliferation and verify drug resistance. The mitochondrial morphology was observed by transmission electron microscopy (TEM) and immunofluorescence staining. The expressions of SIRT3 and OPA1 were detected by real‑time fluorescent quantitative polymerase chain reaction (RTFQ‑ PCR) and Western blot. JC-1 staining was used to detect mitochondrial membrane potential, and dihydroethidium (DHE) staining was used to detect reactive oxygen species (ROS) to verify mitochondrial function. SIRT3 was knocked down in drug-resistant cells by RNA interference, and SIRT3 and YME1L1 wild type (WT), simulated acetylation state mutant (MUT K237Q), and simulated deacetylation state mutant (MUT K237R) were overexpressed in parental cells by overexpression plasmid. Immunoprecipitation assay (IP) and immunofluorescence (IF) were used to verify the interaction between SIRT3 and YME1L1. Results: RTFQ-PCR and Western blot results showed that SIRT3 gene expression and protein level was significantly higher in drug‑resistant cells than in parental cells. Overexpression of SIRT3 in parental cells decreased the sensitivity of breast cancer cells to tamoxifen. Knockdown of SIRT3 in drug-resistant cells enhanced the sensitivity of drug-resistant cells to tamoxifen. DHE staining showed that the ROS level was lower in tamoxifen resistant cells than in parental cells at the same concentration. Transmission electron microscopy and fluorescence staining showed that the mitochondria of the drug-resistant cells were elongated compared with the parental cells. Western blot results showed that the expression level of L-OPA1 protein was higher in drug-resistant cells than in parental cells. Overexpression of SIRT3 in the parental cells resulted in enhanced mitochondrial function and longer mitochondrial morphology compared with the control cells. Western blot showed that the expression of L-OPA1 was upregulated. When SIRT3 was knocked down in drug-resistant cells, the opposite result was obtained. We further verified how SIRT3 regulated OPA1 protein, affected the morphology and function of mitochondria, and promoted drug resistance of breast cancer. Overexpression of YME1L1 (wild-type and mutant plasmids) in parental cells showed that overexpression of YME1L1 in the simulated deacetylation state resulted in similar results as overexpression of SIRT3, and overexpression of YME1L1 in the acetylated state resulted in similar results as knockdown of SIRT3. IP assay confirmed the interaction between SIRT3 and YME1L1 in breast cancer cells. The acetylation level of YME1L1 was different at different SIRT3 expression levels. IF assay showed that YME1L1 was co‑localized with SIRT3 in MCF‑ 7 cells. Conclusion: SIRT3 is highly expressed in tamoxifen-resistant breast cancer cells. SIRT3 upregulates L-OPA1 expression by deacetylating YME1L1, thereby promoting mitochondrial fusion and enhancing mitochondrial function, and promotes tamoxifen resistance in breast cancer.
[Key words] Breast cancer; Endocrine resistance; SIRT3; YME1L1; OPA1
根据2022年全球癌症统计,女性乳腺癌发病率仍位高居,严重威胁女性的身心健康[1]。尽管肺癌在中国仍旧是死亡最多的癌症类型,但女性乳腺癌的发病率及死亡率仍旧居高不下,甚至逐年上升[2]。自 2007年以来,有部分国家癌症登记数据发现,发病率上升仅限于雌激素受体阳性癌症,而雌激素受体阴性的癌症发病率正在下降[3]。雌激素受体阳性患者预后相对较好,然而大多数患者在经过一段时间的内分泌治疗后会产生耐药(获得性耐药),甚至初始内分泌治疗便不敏感(原发性耐药)[4],因此克服内分泌治疗耐药成为研究的重点问题。
调节蛋白3(sirtuin-3,SIRT3)是sirtuins家族的成员,与肿瘤发生、转移和耐药有关[5]。SIRT3主要存在于线粒体中,是线粒体内重要的去乙酰化修饰酶,能够调控线粒体中许多代谢酶的活性,进而调控细胞线粒体的稳态[6-7]。有研究[8-9]表明在乳腺癌内分泌治疗耐药时SIRT3表达水平升高,但具体分子机制仍需探究。
线粒体质量控制对于细胞稳态至关重要,过度的应激导致线粒体稳态失衡,进而导致能量代谢紊乱,最终导致细胞死亡[10]。最新的研究[11]显示,化疗后残存三阴性乳腺癌细胞的线粒体结构和功能出现适应性转变。融合和分裂稳态是线粒体质量控制中最重要的环节,也被称为线粒体动力学[12]。视神经萎缩相关蛋白1(optic atrophy 1,OPA1)参与线粒体内膜融合,受两种线粒体蛋白酶OMA1和YME1样三磷酸腺苷酶(YME1 like 1 ATPase,YME1L1)的水解和调控,OPA1主要以长型(L-OPA1)和短型(S-OPA1)两种形式存在。L-OPA1主要负责线粒体内膜的融合,维持线粒体的正常功能;相反,S-OPA1的积累有助于线粒体分裂,研究[13]表明,YME1L1是稳定L-OPA1的关键酶。研究[14]发现,OPA1可以抑制三阴性乳腺癌的生长。线粒体的融合分裂与乳腺癌内分泌治疗耐药是否相关,仍不清楚。
本研究旨在探讨乳腺癌SIRT3在乳腺癌中的表达及促进乳腺癌内分泌治疗耐药的作用,以及SIRT3、YME1L1、OPA1之间的相互作用,为克服乳腺癌内分泌治疗耐药寻找新的靶点。
1 资料和方法
1.1 主要试剂
DMEM培养基、胎牛血清(fetal bovine serum,FBS)购自武汉普诺赛生命科技有限公司,二甲基亚砜、他莫昔芬(tamoxifen,TAM)购自美国Sigma-Aldrich公司,细胞计数试剂盒(cell counting kit-8)购自艾普拜生物科技(苏州)有限公司,过表达质粒、siRNA均购自上海吉玛制药技术有限公司,LipofectamineTM3000转染试剂购自美国Invitrogen 公司,反转录试剂盒购自日本TaKaRa公司,实时荧光定量聚合酶链反应(real‑time fluorescence quantitative polymerase chain reaction,RTFQ-PCR)试剂购自北京全式金生物技术有限公司,JC-1染色、免疫沉淀(immunoprecipitation,IP)试剂盒购自上海碧云天生物技术股份有限公司,Hoechst 33342染色液购自北京索莱宝科技有限公司,活性氧检测试剂二氢乙啶(dihydroethidium,DHE)、4‘,6-二脒基-2-苯基吲哚(4‘,6-diamidino-2-phenylindole,DAPI)购自北京普利莱基因技术有限公司,MitoTrackerTMRed CMXRos购自美国Thermo Fisher公司,抗SIRT3、抗YME1L1、抗OPA1和抗GAPDH均购自美国Proteintech公司,免疫荧光一抗SIRT3抗体购自美国Santa Cruz公司,抗乙酰化赖氨酸、羊抗鼠Alexa Fluor488、羊抗鼠Alexa Fluor647荧光二抗和酶标结合的二抗购自美国Signalway Antibody LLC公司。
1.2 细胞培养及转染
人乳腺癌细胞(MCF-7)购自厦门逸漠生物科技有限公司。细胞在含有10%FBS和1%青霉素/链霉素的高糖DMEM培养基中于37 ℃、CO2体积分数为5%的条件下培养。MCF-7/TAM细胞在持续存在1 μmol/L TAM的高糖DMEM培养基中培养,添加10%FBS、1%青霉素/链霉素。实验组用TAM为4-羟他莫昔芬(4-hydroxytamoxifen)(H6278,10 μmol/L)刺激48 h。将生长状态良好的对数期细胞铺于6孔板,用脂质体转染法将过表达质粒、siRNA等转入乳腺癌细胞中。操作过程均使用无酶耗材,严格按照 LipofectammeTM3000 使用说明进行转染,并检测转染效率。
1.3 蛋白质印迹法(Western blot)检测
细胞在含有蛋白酶抑制剂和苯甲磺酰氟(phenylmethylsulfonyl-fluoride,PMSF)的蛋白提取缓冲液中裂解,并在4 ℃下以13 000 r/min离心5 min。用二喹啉甲酸(bicinchoninic acid,BCA)蛋白测定试剂盒(北京普利莱基因技术有限公司)定量上清液的蛋白浓度,与上样缓冲液混合并在100 ℃煮沸5 min。样品经十二烷基硫酸钠聚丙烯酰胺凝胶电泳(sodium dodecylsulphate polyacrylamide gel electrophoresis,SDS-PAGE)分离后转移至聚偏二氟乙烯(polyvinylidene fluoride,PVDF)膜上。将膜在室温下用5%的脱脂奶粉封闭2 h,并与一抗(SIRT3 1∶4 000,OPA1 1∶4 000,YME1L1 1∶2 000,GAPDH 1∶1 000)4 ℃温育过夜。将HRP标记的二抗加到膜上并温育22 h,然后用ECL发光液(苏州莫纳生物科技有限公司)和Western blot 曝光仪器显影。使用image J 8.0 软件进行灰度值分析。
1.4 RTFQ-PCR检测
使用TRIzol法提取细胞RNA,酶标仪检测其纯度和浓度。按照转录试剂盒说明书,在 37 ℃ 15 min,85 ℃ 5 s,4 ℃反转录条件下合成cDNA。将各引物和合成的cDNA 按照 PCR 试剂盒说明书体系配置混合液,设置反应条件:预变性95 ℃ 30 s;循环反应为40个循环,95 ℃ 10 s,60 ℃ 30 s;融解曲线95 ℃ 15 s, 60 ℃ 60 s,95 ℃ 15 s。引物内参GAPDH的上游引物序列为5’-ACCCAGAAGACTGTGGAT GG-3’,下游引物序列为5’-TCAGCTCAGGGATG ACCTTG-3’。引物SIRT3的上游引物序列为5’-CCCAGTGGCATTCCAGACTT-3’,下游引物序列为5’-AAGGGCTTGGGGTTGTGAAA-3’。用2-ΔΔCt法计算SIRT3的相对表达量。
1.5 细胞计数试剂盒(cell counting kit-8,CCK-8)检测
乳腺癌细胞转染24~48 h,消化并重新铺板于96孔板中0(每孔5 000 个细胞,100 μL);待细胞贴壁后更换含有不同浓度得TAM的培养基,24 h后更换含有CCK-8工作液浓度的培养基,每孔100 μL,加入过程注意避光,室温温育2 h上机测定 450 nm 处吸光度(D)值。
1.6 透射电镜
用细胞刮刀刮取细胞并用戊二醛和饿酸固定。经不同浓度的乙醇脱水后再次渗透及包埋,使用超薄切片机切片(德国Leica公司),切片经过醋酸铀及枸橼酸铅染色,使用透射电镜观察线粒体形态(日本日立公司)。所得结果采用Image J 8.0 软件进行线粒体形态分析。
1.7 线粒体膜电位测定
采用JC-1试剂盒检测MCF-7及转染后,经TAM处理前后线粒体膜电位的变化。将JC-1染料加入到处理过的细胞中,在37 ℃下温育20 min。用检测缓冲液清洗细胞2次,并将细胞置于培养基中,然后用荧光显微镜捕获。
1.8 IP实验
IP实验按照试剂盒说明书进行。在适当的处理后,从MCF-7细胞中提取蛋白质,并与所指示的一抗(乙酰化赖氨酸或SIRT3)和蛋白质A+G琼脂糖悬浮珠在4 ℃下温育过夜。将磁珠离心洗涤3次,然后将免疫复合物在上样缓冲液中煮沸洗脱。为了确定YME1L1的乙酰化程度以及SIRT3和YME1L1之间的相互作用,蛋白质通过SDS-PAGE分离,并与所指示的一抗(YME1L1 1∶3 000)在4 ℃下温育过夜。其他步骤与Western blot的方法相同。
1.9 线粒体蛋白提取
各组细胞刺激结束后,用细胞刮刀刮取细胞,收集细胞悬液于Eppendorf试管中。根据说明书,使用线粒体分离试剂盒(美国Thermo Scientific公司)提取线粒体蛋白。
1.10 免疫荧光检测
将细胞(3×105个)培养在共聚焦皿内,进行转染等处理。4%多聚甲醛在室温下固定15 min,0.5%的Triton100通透20 min。2%牛血清白蛋白(bovine serum albumin,BSA)封闭 30 min,温育1抗(1∶400)于4 ℃过夜。室温避光温育荧光二抗(1∶100)1 h,洗涤3次。DAPI染色5 min。对于线粒体进行免疫荧光(immunofluorescence,IF)染色,将MitoTrackerTMRed CMXRos和Hoechst 33342染色液配制成工作液,共同温育40 min。用共聚焦显微镜采集图像。
1.11 活性氧的检测
采用荧光探针DHE检测乳腺癌细胞中活性氧(reactive oxygen species,ROS)水平。将细胞(3×105个)培养在共聚焦皿内细胞处理后,DHE配制成工作液温育40 min,DAPI染色 5 min,共聚焦显微镜采集图像。
1.12 统计学处理
采用Graphpad Prism 9.0对数据进行统计学分析。符合正态分布的数据以x±s表示,多组数据间采用单因素方差分析,两组之间数据分析采用t检验。P<0.05为差异有统计学意义。
2 结 果
2.1 构建稳定的乳腺癌耐药细胞株
本研究采用TAM冲击法,诱导人乳腺癌MCF-7/TAM耐药细胞株。后续以浓度为1 μmol/L TAM的培养基维持耐药性。结果显示,耐药组的细胞在不同浓度的TAM下细胞成活率更高,证明我们成功构建出耐药株(P<0.01,图1)。
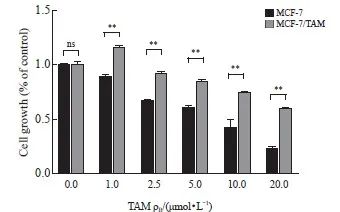
图1 构建稳定的乳腺癌耐药细胞株
Fig. 1 Construction of stable drug-resistant breast cancer cell lines
CCK-8 assay was used to detect the cell viability under tamoxifen treatment. **: P<0.01, MCF-7 compared with MCF-7/TAM. ns: No significance.
2.2 SIRT3在乳腺癌耐药细胞中高表达
检测乳腺癌亲本细胞及耐药细胞株中SIRT3的表达情况,通过RTFQ-PCR以及Western blot检测乳腺癌细胞(MCF-7)、耐药细胞系(MCF-7/TAM)细胞中SIRT3的表达水平。在两种乳腺癌细胞中,耐药细胞中SIRT3的RNA及蛋白表达水平高于亲本乳腺癌细胞系(P<0.05,图2)。
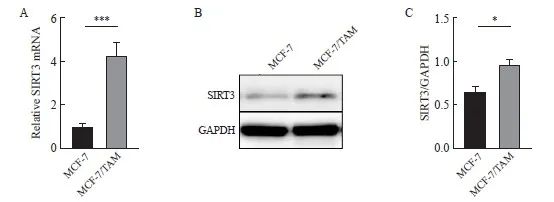
图2 SIRT3在TAM耐药的乳腺癌中高表达。
Fig. 2 SIRT3 is highly expressed in tamoxifen-resistant breast cancer.
A: SIRT3 mRNA expression was detected by RTFQ‑PCR. B, C: SIRT3 protein expression and quantification in both cell lines were detected by Western blot. *: P<0.05, compared with each other; ***: P<0.001, compared with each other.
2.3 乳腺癌细胞耐药后,线粒体动力学和功能发生改变
线粒体荧光探针观察耐药组线粒体形态相对更长(P<0.01,图3A、B),提示乳腺癌细胞耐药后线粒体融合增加,分裂减少。电镜观察结果显示,耐药细胞线粒体多呈长管状,亲本细胞线粒体呈圆形、短小,线粒体纵横比减小(P<0.01,图3C、D)。Western blot检测结果显示,L-OPA1表达水平在耐药后上调(P<0.05,图3E、F)。使用ROS特异性荧光探针DHE评估氧化应激水平和线粒体功能。结果显示,在对TAM的刺激下,耐药MCF-7/TAM细胞产生的ROS较亲本MCF-7的水平明显降低。这些结果表明TAM耐药导致线粒体功能增强(P<0.01,图3G、H)。
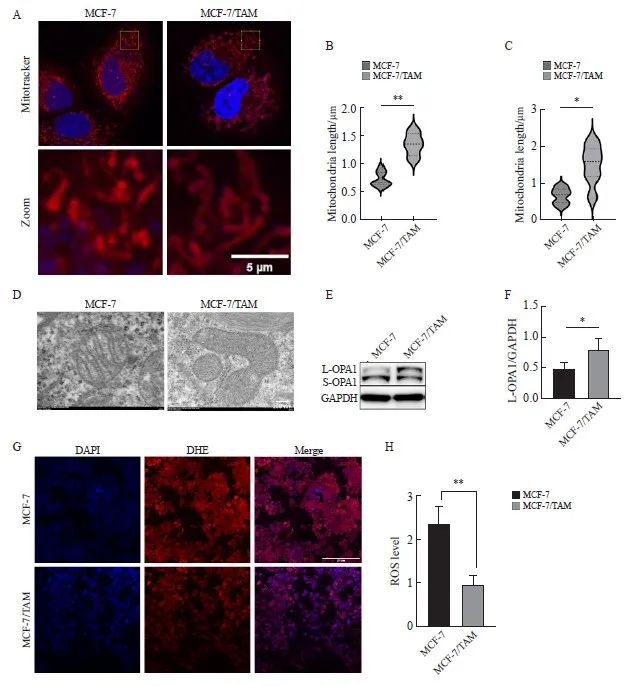
图3 乳腺癌内分泌耐药后,检测线粒体形态及功能
Fig. 3 Detection of mitochondrial morphology and function in breast cancer after endocrine resistance
A, B: Mitochondrial length was observed and quantified by fluorescence staining (scale bar=5 μm); C, D: Mitochondrial morphology was observed and quantified by transmission electron microscopy (scale bar=200 nm); E, F: Western blot was used to detect and quantify L-OPA1 protein in parental cells and drug‑resistant cells. G, H: DHE staining assessment, ROS levels and quantification in parental and resistant cells (scale bar=200 μm). *: P<0.05, compared with each other; **: P<0.01, compared with each other.
2.4 敲低SIRT3促进MCF-7/TAM对药物敏感
为了进一步研究SIRT3在耐药细胞中的作用,在耐药细胞中使用si-RNA敲低SIRT3。结果显示,相较于对照组,敲低组SIRT3的表达水平显著降低(P<0.05,图4A、B)。CCK-8实验检测TAM处理下细胞的活力,结果表明,相较于对照组,SIRT3敲低组显著增加了耐药细胞对TAM的敏感性(P<0.05,图4D)。IF观察到敲低SIRT3加剧了线粒体形态的改变,线粒体长度较对照组明显缩短(P<0.01,图4F、H)。Western blot结果显示,在敲低组,L-OPA1表达降低(P<0.05,图4A、C),进一步支持了IF染色线粒体发现的线粒体形态改变的结果。TAM诱导的ROS水平通过活性氧检测(DHE探针),结果显示,相较于对照组,敲低组的活性氧水平明显升高(P<0.01,图4E、G)。
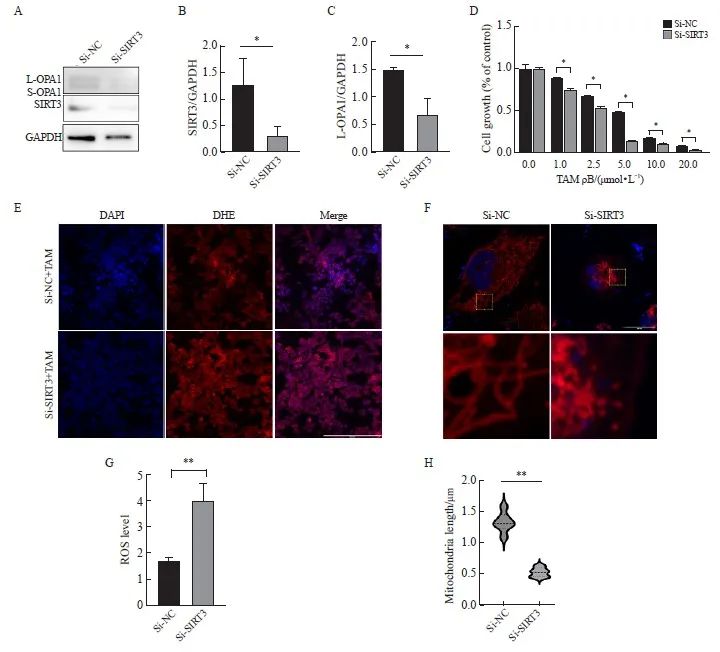
图4 在耐药细胞中敲低SIRT3检测细胞增殖、线粒体形态及功能
Fig. 4 Cell proliferation, mitochondrial morphology, and function were examined by SIRT3 knockdown in drug-resistant cells
A, B, C: Western blot was used to detect the expression and quantification of SIRT3 protein and L-OPA1 protein after SIRT3 knockdown. D: CCK-8 assay was used to detect the cell viability under tamoxifen treatment; E, G: DHE probe was used to detect the intracellular ROS level and quantification after SIRT3 knockdown (scale bar=200 μm); F, H: Immunofluorescence detection of mitochondrial length changes and quantification after SIRT3 knockdown (scale bar=5 μm). *: P<0.05, Si-NC compared with Si-SIRT3.
2.5 过表达SIRT3促进MCF-7(亲本细胞)对TAM耐药
在亲本细胞MCF-7细胞中,我们使用质粒过表达SIRT3。结果显示,相较于对照组,SIRT3过表达组的表达水平升高(P<0.05,图5A、B)。CCK-8实验检测TAM处理下细胞的活力。结果表明,相较于对照组,SIRT3过表达显著增加了细胞对TAM的耐药性(P<0.05,图5D)。线粒体膜电位检测,结果显示,在TAM处理下,相较于对照组,过表达SIRT3组的线粒体膜电位更高(P<0.01,图5E、G)。IF观察到过表达SIRT3后,线粒体长度较对照组明显变长(图5H、I)。Western blot结果显示,在过表达组,L-OPA1表达升高(P<0.01,图5A、C)。
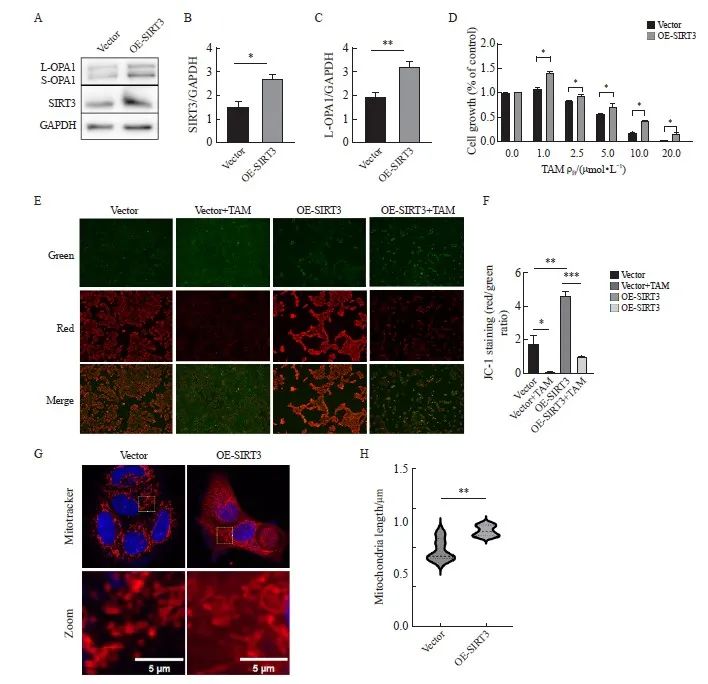
图5 在亲本细胞中过表达SIRT3检测细胞增殖、线粒体形态及功能
Fig. 5 Overexpression of SIRT3 in parental cells examined cell proliferation, mitochondrial morphology, and function
A, B, C: Western blot was used to detect the expression and quantification of L‑OPA1 protein after SIRT3 overexpression; D: After SIRT3 was overexpressed in parental cells, CCK‑8 assay was used to detect the cell viability under tamoxifen treatment; E, G: Detection and quantification of mitochondrial membrane potential by JC-1 probe (scale bar=200 μm); H, I: Fluorescence staining for mitochondrial morphology and quantification (scale bar=5 μm). *: P<0.05, compared with each other; **: P<0.01, compared with each other; ***: P<0.001, compared with OE-SIRT3 group.
2.6 YME1L1的乙酰化调控OPA1介导的线粒体融合
本研究推测SIRT3可能通过作用于YME1L1调控OPA1,进而影响线粒体动力学。通过Uniprot数据库查询YME1L1的氨基酸序列,发现YME1L1人亚型1的第237位有一个赖氨酸(K)位点。在MCF-7细胞中我们转染位点突变的YME1L1质粒,以模拟YME1L1在细胞内不同乙酰化状态。YME1L1-K237Q质粒将赖氨酸237(K)突变为谷氨酰胺(Q)以模拟乙酰化状态。YME1L1-K237R质粒将赖氨酸237(K)突变为精氨酸(R)以模拟去乙酰化状态。Western blot结果显示,与转染YME1L1-WT相比,转染YME1L1-K237R质粒,L-OPA1蛋白表达升高,转染YME1L1-K237Q质粒,L-OPA1蛋白表达降低(P<0.01,图6A、B)。与转染YME1L1-WT组相比,转染YME1L1-K237Q质粒组的线粒体分裂更多,转染YME1L1-K237R质粒组线粒体融合更多(P<0.01,图6C、D)。
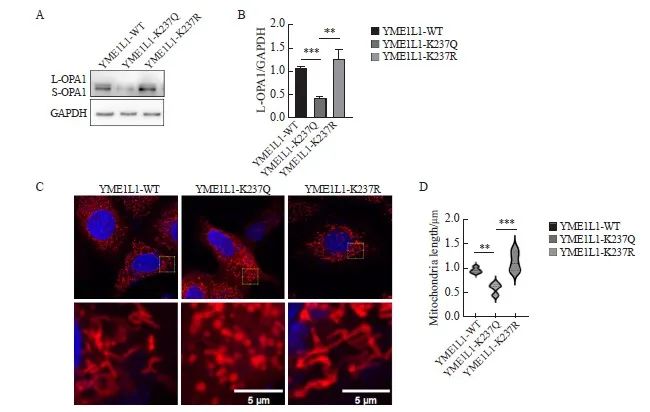
图6 在亲本细胞中过表达YME1L1导致线粒体形态改变
Fig. 6 Changes in mitochondrial morphology caused by overexpression of YME1L1 in parental cells
A, B: YME1L1 was overexpressed, and the expression and quantification of L‑OPA1 protein were detected by Western blot. C, D: Length and quantification of mitochondria by fluorescent staining (scale bar=5 μm). **: P<0.01, compared with each other; ***: P<0.001, compared with YME1L1-K237Q.
2.7 SIRT3通过去乙酰化YME1L1作用于OPA1
在MCF-7细胞中过表达SIRT3,在MCF-7/TAM细胞中敲低SIRT3。IP结果显示,与Ac-K抗体结合的蛋白中检测到YME1L1蛋白,且相较于对照组,过表达SIRT3组中的YME1L1蛋白较少(图7A~C);在敲低SIRT3组中得到相反结果(P<0.05,图7D、E)。通过IP实验进一步验证SIRT3与YME1L1具有相互结合作用,结果显示,在与SIRT3抗体结合的蛋白中能够检测到YME1L1蛋白(图7G)。IF结果也显示,在乳腺癌细胞中,SIRT3与YME1L1存在相互作用(图7H)。
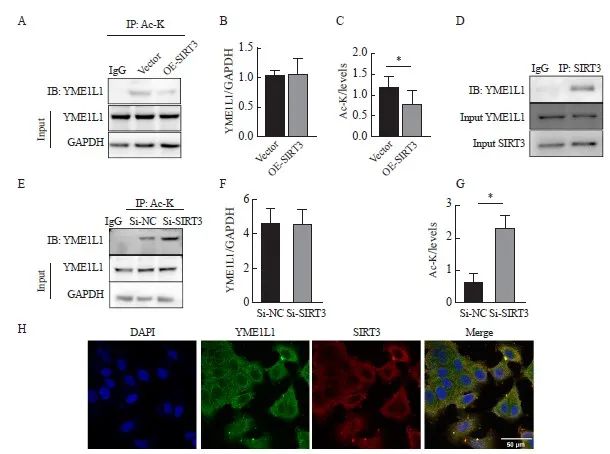
图7 SIRT3调控YME1L1的乙酰化状态
Fig. 7 SIRT3 regulates the acetylation status of YME1L1
A-G: IP was used to detect the interaction between YME1L1 and SIRT3 and quantify. *: P<0.05, compared with each other; **: P<0.01, compared with each other; ***: P<0.001, compared with each other. H: Co‑localization of YME1L1 and SIRT3 was detected by immunofluorescence (scale bar=50 μm).
2.8 去乙酰化YME1L1促进乳腺癌对TAM耐药
在亲本细胞中导入模拟不同乙酰化状态的YME1L1质粒后,CCK-8结果显示,与野生型(YME1L1-WT)组相比,乙酰化(YME1L1-K237Q)组的耐药性下降,去乙酰化(YME1L1-K237R)组的耐药性增强(P<0.05,图8)。
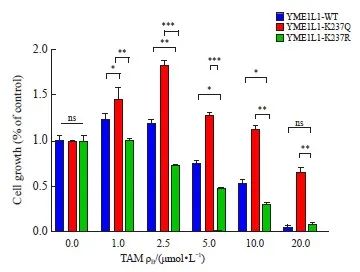
图8 去乙酰化YME1L1促使乳腺癌对TAM耐药
Fig. 8 Deacetylating Y ME1L1 promotes tamoxifen resistance in breast cancer
CCK-8 assay was used to detect cell viability under tamoxifen treatment. *: P<0.05, compared with each other; **: P<0.01, compared with each other; ***: P<0.001, compared with each other. ns: No significance, compared with YME1L1‑WT or YME1L1‑K237R.
3 讨 论
耐药的出现限制了患者的治疗效果[15]。SIRT3作为一种主要的线粒体NAD+依赖性去乙酰化酶,在应对各种应激时对线粒体代谢适应至关重要[16]。SIRT3与肿瘤发生、转移、耐药等密切相关[17-18]。SIRT3在不同癌症中的作用不同,并且具有特异性;即使在相同的癌症中,不同研究人员也有不同的观点。有研究[19]表明, SIRT3在乳腺癌中发挥致癌作用。例如,在同一位乳腺癌患者的肿瘤组织中,SIRT3阳性的乳腺癌细胞通过激活SIRT/FOXO/SOD2轴上调线粒体未折叠蛋白(UPRmt)从而表现出更强的侵袭性。乳腺癌患者淋巴结转移,也与SIRT3的表达呈正相关[20]。一项研究[21]表明,SIRT3对脯氨酸5-羧基还原酶1(PYCR1)在K228位点的去乙酰化作用可以调节脯氨酸代谢,并促进MCF-7细胞的生长。然而也有研究[22]表明SIRT3在乳腺癌中表现出抑制作用。例如,SIRT3的基因缺失会导致异常的线粒体结构、基因组结构不稳定以及应激诱导超氧化物增加,敲除SIRT3基因的小鼠可自发形成乳腺肿瘤。另一项研究[23]发现,SIRT3低水平表达的乳腺癌患者的无复发生存期相对较短。Finley等[24]发现乳腺癌中SIRT3表达降低,过表达SIRT3可以抑制乳腺癌细胞中的糖酵解和增殖。本研究发现,在雌激素受体阳性乳腺癌细胞中SIRT3呈高表达,可促使癌细胞对TAM敏感性下降。
肿瘤细胞的氧化应激水平增加及细胞凋亡异常与线粒体相关,因此线粒体功能的调节与癌症的发生、发展密切相关。线粒体具有高度动态的结构,在活细胞内经历或长或短的形态学变化,这种高度调节的过程被称为线粒体动力学[25]。YME1L1是ATP酶AAA家族的主要成员,对于维持线粒体形态、功能和可塑性至关重要[26]。OPA1的功能受YME1L和OMA1肽酶影响,生成一种与线粒体膜结合的长型L-OPA1,或另一种可溶的短型S-OPA1[27]。有研究[28]报道,OPA1蛋白在肿瘤血管生成过程中扮演重要角色。在乳腺癌细胞中,抑制OPA1可以增加miR-148/152的水平,降低乳腺癌的迁移、侵袭能力[14];化疗后残存三阴性乳腺癌细胞通过OPA1介导线粒体融合,增加OXPHOS能力,从而对化疗出现适应[11]。本研究发现,在雌激素受体阳性乳腺癌中导入YME1L1突变质粒后,L-OPA1蛋白表达水平发生改变,得到与敲降SIRT3相似的结果,乳腺癌细胞对TAM出现耐药。
综上所述,SIRT3蛋白在乳腺癌内分泌耐药后表达上调,SIRT3通过使YME1L1去乙酰化作用于OPA1蛋白,促进线粒体融合,使乳腺癌对TAM产生耐药性。SIRT3-YME1L1-OPA1信号转导通路可能是乳腺癌内分泌治疗耐药的重要因素,本研究结果可望为乳腺癌内分泌治疗耐药提供新的思路。
利益冲突声明:所有作者均声明不存在利益冲突。
作者贡献声明:董涧桥负责文章撰写,实施研究过程;李坤艳负责数据分析;李菁负责实验指导,图片整理;王斌负责文献整理,文章修改;王艳红负责实验监督,文章修改;贾红燕负责设计研究思路及方案,审核文章。
[参考文献]
[1] BRAY F, LAVERSANNE M, SUNG H, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA A Cancer J Clin, 2024, 74(3): 229-263.
[2] CAO W, CHEN H D, YU Y W, et al. Changing profiles of cancer burden worldwide and in China: a secondary analysis of the global cancer statistics 2020[J]. Chin Med J, 2021, 134(7): 783-791.
[3] SIEGEL R L, GIAQUINTO A N, JEMAL A. Cancer statistics, 2024[J]. CA Cancer J Clin, 2024, 74(1): 12-49.
[4] VOUDOURI K, BERDIAKI A, TZARDI M, et al. Insulin-like growth factor and epidermal growth factor signaling in breast cancer cell growth: focus on endocrine resistant disease[J]. Anal Cell Pathol, 2015, 2015: 975495.
[5] ZAHEDIPOUR F, JAMIALAHMADI K, KARIMI G. The role of noncoding RNAs and sirtuins in cancer drug resistance[J]. Eur J Pharmacol, 2020, 877: 173094.
[6] LIU F, YUAN L H, LI L, et al. S-sulfhydration of SIRT3 combats BMSC senescence and ameliorates osteoporosis via stabilizing heterochromatic and mitochondrial homeostasis[J]. Pharmacol Res, 2023, 192: 106788.
[7] ZHANG J, XIANG H G, LIU J, et al. Mitochondrial Sirtuin 3: new emerging biological function and therapeutic target[J]. Theranostics, 2020, 10(18): 8315-8342.
[8] ZHANG L, REN X C, CHENG Y, et al. Identification of Sirtuin 3, a mitochondrial protein deacetylase, as a new contributor to tamoxifen resistance in breast cancer cells[J]. Biochem Pharmacol, 2013, 86(6): 726-733.
[9] HUBER-KEENER K J, LIU X P, WANG Z, et al. Differential gene expression in tamoxifen-resistant breast cancer cells revealed by a new analytical model of RNA-Seq data[J]. PLoS One, 2012, 7(7): e41333.
[10] Disturbance of mitochondrial dynamics and mitophagy in sepsis-induced acute kidney injury-PubMed [Internet].[cited 2024 Mar 2]. https://pubmed.ncbi.nlm.nih.gov/31479679/.
[11] BAEK M L, LEE J, PENDLETON K E, et al. Mitochondrial structure and function adaptation in residual triple negative breast cancer cells surviving chemotherapy treatment[J]. Oncogene, 2023, 42(14): 1117-1131.
[12] ADEBAYO M, SINGH S, SINGH A P, et al. Mitochondrial fusion and fission: the fine-tune balance for cellular homeostasis[J]. FASEB J, 2021, 35(6): e21620.
[13] RUAN Y, LI H, ZHANG K, et al. Loss of Yme1L perturbates mitochondrial dynamics[J]. Cell Death Dis, 2013, 4(10): e896.
[14] ZAMBERLAN M, BOECKX A, MULLER F, et al. Inhibition of the mitochondrial protein Opa1 curtails breast cancer growth[J]. J Exp Clin Cancer Res, 2022, 41(1): 95.
[15] HANKER A B, SUDHAN D R, ARTEAGA C L. Overcoming endocrine resistance in breast cancer[J]. Cancer Cell, 2020, 37(4): 496-513.
[16] CARRICO C, MEYER J G, HE W J, et al. The mitochondrial acylome emerges: proteomics, regulation by sirtuins, and metabolic and disease implications[J]. Cell Metab, 2018, 27(3): 497-512.
[17] LIU L G, LI Y, CAO D Y, et al. SIRT3 inhibits gallbladder cancer by induction of AKT-dependent ferroptosis and blockade of epithelial-mesenchymal transition[J]. Cancer Lett, 2021, 510: 93-104.
[18] S O, Q Z, L L, K Z, Z L, P L, et al. The double-edged sword of SIRT3 in cancer and its therapeutic applications[J]. Front Pharmacol, 2022, 13: 871560.
[19] KENNY T C, HART P, RAGAZZI M, et al. Selected mitochondrial DNA landscapes activate the SIRT3 axis of the UPRmt to promote metastasis[J]. Oncogene, 2017, 36(31): 4393-4404.
[20] ASHRAF N, ZINO S, MACINTYRE A, et al. Altered sirtuin expression is associated with node-positive breast cancer[J]. Br J Cancer, 2006, 95(8): 1056-1061.
[21] CHEN S Y, YANG X, YU M, et al. SIRT3 regulates cancer cell proliferation through deacetylation of PYCR1 in proline metabolism[J]. Neoplasia, 2019, 21(7): 665-675.
[22] KIM H S, PATEL K, MULDOON-JACOBS K, et al. SIRT3 is a mitochondria-localized tumor suppressor required for maintenance of mitochondrial integrity and metabolism during stress[J]. Cancer Cell, 2010, 17(1): 41-52.
[23] DESOUKI M M, DOUBINSKAIA I, GIUS D, et al. Decreased mitochondrial SIRT3 expression is a potential molecular biomarker associated with poor outcome in breast cancer[J]. Hum Pathol, 2014, 45(5): 1071-1077.
[24] FINLEY L W S, CARRACEDO A, LEE J, et al. SIRT3 opposes reprogramming of cancer cell metabolism through HIF1α destabilization[J]. Cancer Cell, 2011, 19(3): 416-428.
[25] ANDREWS R M, KUBACKA I, CHINNERY P F, et al. Reanalysis and revision of the Cambridge reference sequence for human mitochondrial DNA[J]. Nat Genet, 1999, 23(2): 147.
[26] OHBA Y, MACVICAR T, LANGER T. Regulation of mitochondrial plasticity by the i-AAA protease YME1L[J]. Biol Chem, 2020, 401(6/7): 877-890.
[27] ANAND R, WAI T, BAKER M J, et al. The i-AAA protease YME1L and OMA1 cleave OPA1 to balance mitochondrial fusion and fission[J]. J Cell Biol, 2014, 204(6): 919-929.
[28] HERKENNE S, EK O, ZAMBERLAN M, et al. Developmental and tumor angiogenesis requires the mitochondria-shaping protein Opa1[J]. Cell Metab, 2020, 31(5): 987-1003.e8.

