Diabetologia:膳食棕榈酸酯和油酸酯不同程度地调节人体骨骼肌胰岛素敏感性。
时间:2021-11-14 13:02:48 热度:37.1℃ 作者:网络

西式饮食的定义是摄入高能量和脂肪,这与肥胖和2型糖尿病等胰岛素抵抗状态患病率的上升有关。尽管目前的指南建议2型糖尿病和心血管疾病食用低饱和脂肪酸(FA),但证据仍然有限。以前,我们已经证明肥胖和2型糖尿病的骨骼肌胰岛素抵抗与肌细胞二酰甘油-新的蛋白激酶C亚型θ通路的激活有关,这是近端胰岛素信号转导的一个抑制级联反应。与临床前模型的结果一致,我们发现该通路在葡萄糖耐受性人群静脉输注含主要单不饱和脂肪酸的脂乳时也起作用,在肥胖个体和不输注的2型糖尿病患者中也起作用。值得注意的是,这项研究没有检测到其他可能导致胰岛素抵抗的机制的变化,例如鞘磷脂介质、线粒体功能异常或低度炎症。在其他原因中,NEFA饱和的程度可能会导致不同的结果,这可能会不同地影响新陈代谢和2型糖尿病和心血管疾病的风险,因为饱和FA,而不是单一不饱和FA,被认为是有害的。已知单不饱和脂肪酸可激活DAG-nPKCAkt途径,而饱和脂肪酸可增加细胞内神经酰胺,刺激蛋白磷酸酶2A(PP2A)和非典型蛋白激酶C(APKC)亚型ζ,从而抑制Akt的激活。另一方面,油酸甚至可以预防棕榈酸诱导的胰岛素抵抗,就像L6肌管所证明的那样。然而,人体骨骼肌在摄入不同组成的脂类时的分子事件的时间顺序仍不清楚。我们的目的是比较饱和脂肪(棕榈油[PAL])和单不饱和脂肪(红花油[SAF])与水(载体[VCL])对年轻、苗条和新陈代谢健康的人的下列变量的急性影响(ESM表1):(1)全身胰岛素敏感性;(2)脂质中间体的亚细胞分布;(3)胰岛素信号;以及(4)骨骼肌中的线粒体氧化能力。因此,我们分别在干预前后2小时、4小时和7小时进行了一系列活组织检查,进行了全面的代谢表型分析(图1和ESM图1)。
目的:高能量营养通常会导致胰岛素抵抗,但饮食结构可能会不同地影响葡萄糖代谢。本研究调查了单不饱和脂餐与饱和脂餐对基础和胰岛素刺激的心肌细胞葡萄糖代谢和胰岛素信号的初步影响。
方法:所有参与者(10名男性,6名女性)在登记参加这项随机、安慰剂对照的交叉试验之前都给予了书面知情同意。纳入标准为年龄20~40岁,体重指数20~25 kg/m2。诊断标准包括糖尿病家族史、血糖异常、月经不调、吸烟、酗酒或药物滥用史以及其他急性或慢性疾病(包括癌症),以及任何影响胰岛素敏感性、免疫系统或脂代谢的药物摄入。所有志愿者都接受了病史和临床检查、体重评估、常规实验室检查和75g口服葡萄糖耐量试验(OGTT)的筛查。符合条件的参与者被指示保持并记录富含碳水化合物的饮食,并在所有研究日之前的3天内避免剧烈的体力活动。女性参与者只在月经周期的第5天到第9天进行检查。所有参与者被随机分配到三个研究日,间隔3周(图1)。在一项随机交叉研究中,16名瘦肉型代谢健康志愿者接受了含有红花油(SAF)、棕榈油(PAL)或维生素C(VCL)的单餐。从葡萄糖处理(RD)开始和D-[6,6-2H2]葡萄糖钳夹高胰岛素-正常血糖期间测定全身葡萄糖代谢。在连续的骨骼肌活组织检查中,在用餐前后测量亚细胞脂质代谢产物和胰岛素信号。
结果:SAF和PAL均可使血浆油酸浓度升高,但仅PAL能显著升高血浆棕榈酸浓度。SAF和PAL可增加心肌细胞二酰甘油和活化蛋白激酶C亚型θ(p<0.0 5),但仅PAL激活蛋白激酶Cɛ(p<0.0 5)。此外,PAL可导致心肌细胞神经酰胺增加,并刺激蛋白激酶Cζ易位(p<0.0 5 vs SAF)。钳夹期间,SAF和PAL均可降低胰岛素刺激的RD(与VCL相比,p<0.05),但PAL后非氧化葡萄糖处理低于SAF(p<0.05)。肌丝氨酸1101-IRS-1磷酸化在SAF和PAL消耗时增加(p<0.05),而PAL降低Akt的丝氨酸473-磷酸化程度大于SAF(p<0。0 5)。
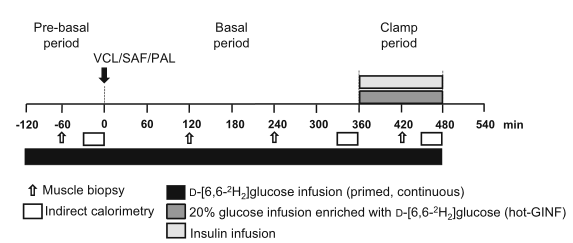
图1 研究设计。健康、体型瘦的成人(男10人,女6人)在12周的时间里,在0分钟的时间点随机摄入一剂PAL、SAF或VCL(水),共三次。从−开始120min,D-[6,6-2H2]葡萄糖输注至+480min。在−60min、+120min、+240min和+420min的时间点进行肌肉活检。从+360分钟到+480分钟,按照热葡萄糖输注(HOT-GINF)方案进行高胰岛素-正常血糖钳夹试验

图2 健康人循环脂质代谢产物的时间进程。血浆乳糜微粒(A)、三酰甘油(B)、总NEFA(C)、棕榈酸(D)、油酸(E)和亚油酸(F)的浓度与PAL(红)、SAF(蓝)或VCL(水、灰)的摄入量成正比。数据显示为均值±扫描电镜;n=16(乳糜微粒n=10)。*p<0.0 5,**p<0.0.001.1和*p<0.0 1与VCL比较,PAL与SAF在同一时间点的†p<0.0 5(对干预之间的每个时间点进行Tukey-Kramer校正,对重复测量进行了ANOVA调整)。
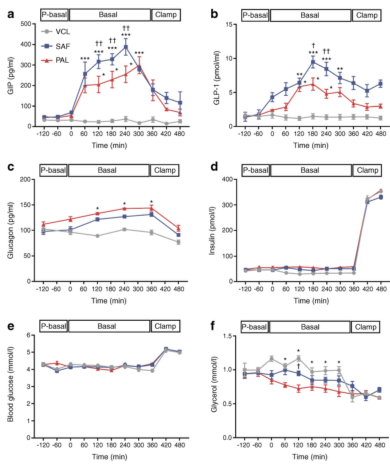
图3 健康人体内循环激素和代谢物的时间进程。在0分钟摄入PAL(红色)、SAF(蓝色)或VCL(水、灰色)后,呈现血浆GIP (a)、血浆GLP-1 (b)、血浆胰高血糖素(c)、血浆胰岛素(d)、血糖(e)和血浆甘油(f)的浓度。数据以扫描电镜的方式显示;胰岛素和血糖n = 16。GIP,GLP-1,胰高血糖素和甘油,n= 4。*p<0.0 5,**p<0.0.001.1和*p<0.0 1与VCL比较,PAL与SAF在同一时间点的†p<0.0 5(对干预之间的每个时间点进行Tukey-Kramer校正,对重复测量进行了ANOVA调整)。
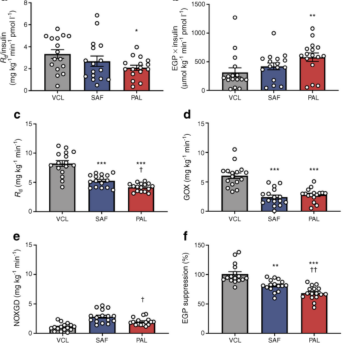
图4 健康人基础期和钳夹期全身葡萄糖处理率。(a,b)在基础期的最后30分钟(+330分钟至+360分钟)内,葡萄糖代谢率与周围血浆胰岛素浓度相关:Rd/胰岛素(a)和EGP×胰岛素(b)。(c–f)在钳夹稳态(+450分钟至+480分钟)期间,在0分钟摄入PAL(红色)、SAF(蓝色)或VCL(水、灰色)后,出现胰岛素刺激的Rd(c)、GOX率(d)、NOXGD率(e)和EGP抑制(f)。数据以扫描电镜的方式显示;n=16。*p<0.0 5,**p<0.0.001.1和*p<0.0 1与VCL比较,PAL与SAF在同一时间点的†p<0.0 5(对干预之间的每个时间点进行Tukey-Kramer校正,对重复测量进行了ANOVA调整)。

图5 健康人的心肌细胞脂质代谢物和胰岛素信号传导(DAG-nPKC途径)。细胞膜组分(a)中的DAG物种18–1:18–1,16–0:16–0和16–0:18–1,脂质液滴组分(b)中的DAG物种18–1:18–1,16–0:16–0 a n D1 6–0:18–1,nPKCε激活(c),nPKCθ激活(d)和IRS-1水平(e)以及IRS-1相对于IRS-1的serine 1101磷酸化针对总蛋白和胞质蛋白的GAPDH和针对膜蛋白的Na+/K+-ATPase进行标准化后,免疫印迹上的表达信号以任意单位(AU)表达。数据以扫描电镜的方式显示;时间点60分钟时n= 16,时间点+120分钟时n= 10,时间点+240分钟和+420分钟时n= 6。*p< 0.05 vs .同一时间点的VCL(对干预之间的每个时间点进行Tukey-Kramer校正,对重复测量进行了ANOVA调整)。LD,脂滴分数
结论:脂质诱导的心肌细胞胰岛素抵抗在棕榈酸盐组可能比油酸盐组更明显,并与PKC亚型激活和胰岛素信号传导抑制有关。
原文出处:
Sarabhai T, Koliaki C, Mastrototaro L,et al.Dietary palmitate and oleate differently modulate insulin sensitivity in human skeletal muscle.Diabetologia 2021 Oct 26

